Illustration of a dreaming brain by Georgia Turner for Cell Press.
A Sleeping Brain Builds a World
Every night, the brain does something that should feel impossible. With the eyes shut, the body motionless, and the external world mostly unavailable, it constructs places, people, threats, conversations, and plots that can feel utterly real while they are happening. The puzzle is not only why dreams are strange, but why a sleeping brain generates a world at all. The answer turns out to matter for far more than sleep science. Dreaming offers a window into something happening in your brain at every moment. Your conscious experience of the world is actually a construction, built moment to moment out of expectations and incoming sensory signals. Dreaming, then, offers a clue to how the brain normally produces this conscious experience because it shows that construction continuing under conditions where the usual rules of perception break down, making that building process visible in a way it never is during normal waking life [1][2].
Scientists have long asked what dreams are for. Some theories treat dreams as rehearsals for threat and danger [3]. Others suggest that sleep helps people rework emotional experiences [4]. Still others focus on memory, since dream content often includes recent events, and dreaming about a task can sometimes predict better later performance on it [5]. These questions matter, but they leave a different question untouched. Even before we know what dreams do, we have to explain how a sleeping brain creates dreams at all. To answer this question, it helps to step back and ask how the brain produces conscious experience in any state, awake or asleep. The predictive processing framework offers exactly this kind of answer, and it places dreaming inside a much larger story of how perception works [2][6][7].
Perception as Controlled Construction
The basic claim of predictive processing sounds simple, but it has strange implications. The brain does not work by passively receiving information from the world. It works by guessing. Imagine walking into your favorite coffee shop. Before you have even crossed the threshold, your brain has already produced a set of expectations. The smell of espresso. The hiss of a steamer. The familiar layout of tables and the rough texture of the wooden counter. These expectations are predictions, generated by higher levels of the brain that handle complex processes like reasoning and abstract thinking, and are sent down to the lower, more primitive level of the brain involving the sensory systems that handle smell, sound, and touch. When the senses confirm the expectation, the brain barely registers anything. When the senses do not match, like if the counter has been replaced with smooth marble overnight, the brain registers the mismatch sharply. That mismatch is called prediction error, representing the gap between what the brain expected and what the senses actually delivered. Prediction error, therefore, is the main currency the brain uses to update its model of the world.
Perception, then, is the brain’s best running guess about what is causing the patterns of light, sound, and pressure hitting the senses, with the senses themselves serving mainly to correct the guess when it goes wrong. Context shapes perception so powerfully because expectation is built into the mechanism rather than added on at the end [6]. Once you accept that perception is built from the inside out, dreams stop seeming so strange. A dream is what happens when the construction process keeps running but the correction process drops away. The brain is still building a world, it just stops checking that world against the senses with the same vigilance.
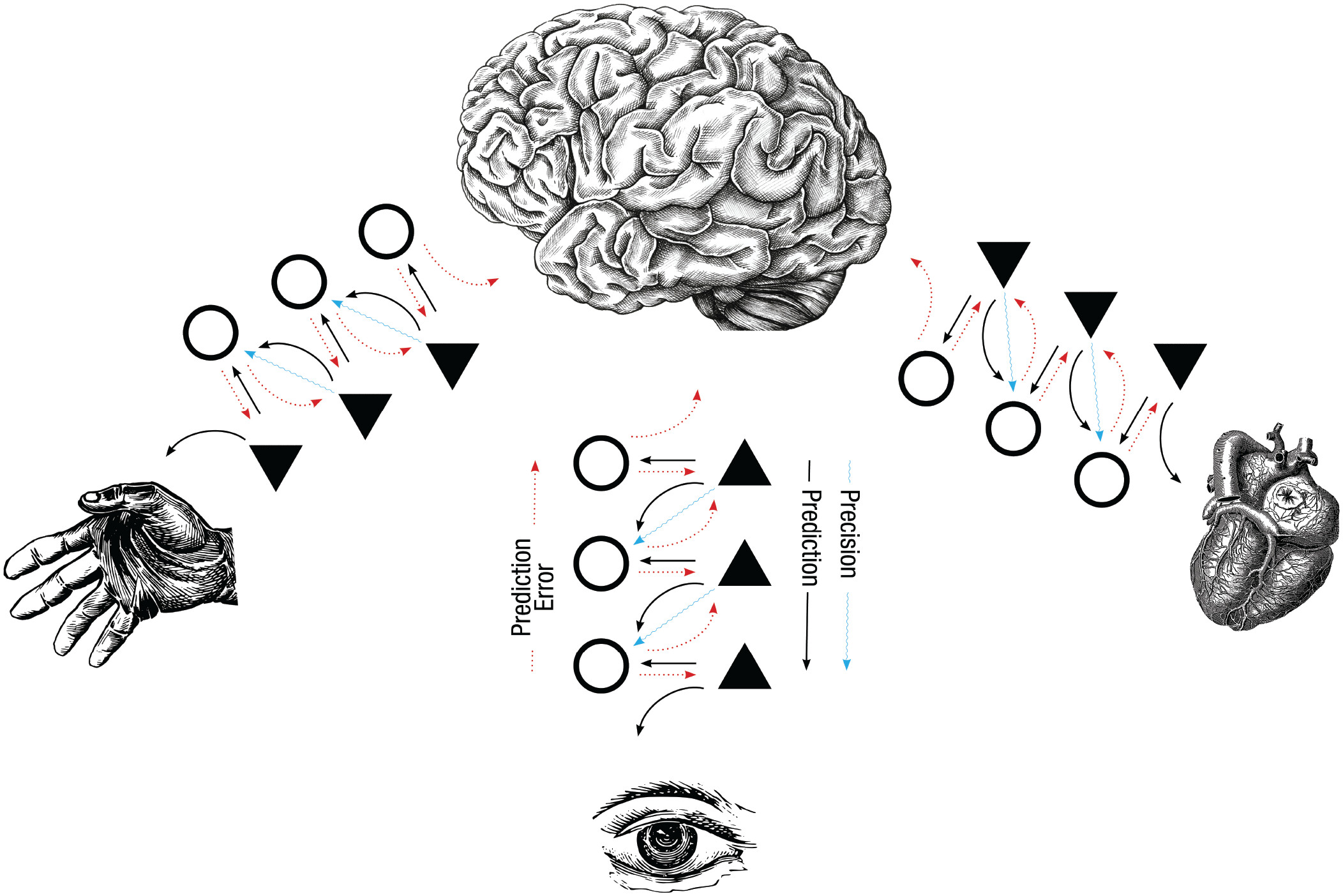
Diagram of the predictive-processing framework. The downward arrows on each side represent predictions flowing from higher brain regions toward sensory systems, including the hand for touch, the eye for vision, and visceral organs for internal sensation. The upward arrows represent prediction errors flowing the other direction, carrying back any mismatch between the prediction and the actual sensory signal. The triangles and circles represent neural populations that handle these two streams. Precision functions like a volume knob, deciding how much weight to give either the prediction or the prediction error in any given moment [8].
Studying a Private World from the Outside
If dreams are internally built worlds, then studying them creates an obvious problem. Researchers cannot directly watch a dream from the outside. Dreams are private, internally generated experiences that fade quickly on waking. Scientists need ways to identify what state the brain is in, wake people at the right moment, and connect the person’s report to measurable brain activity. The history of dream research therefore depends heavily on methods for tracking the sleeping brain.
The most foundational tool arrived in the 1950s with electroencephalography, or EEG, which measures the electrical activity of the brain through small sensors placed on the scalp. EEG can detect changes in brain activity on the scale of milliseconds, which makes it ideal for tracking the rapid shifts that happen during sleep. In a sleep lab, EEG is usually combined with sensors that measure eye movements and muscle tone. The full setup is called polysomnography, and it remains the gold standard for figuring out what stage of sleep someone is in at any given moment [9][10].
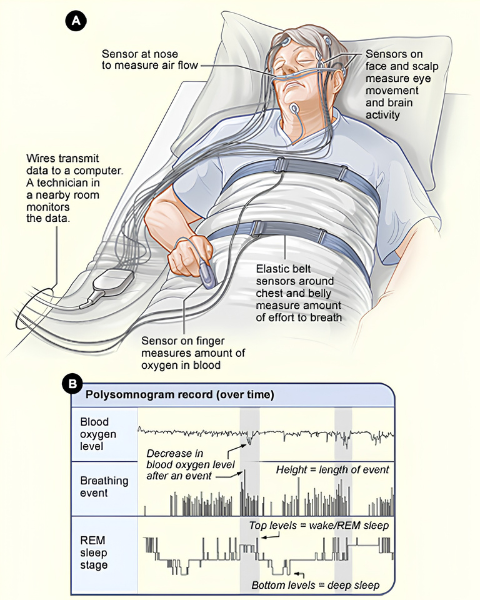
Polysomnography setup and example output. The top panel (A) shows the sensors used during a sleep study. A nose sensor measures airflow, sensors on the face and scalp track eye movements and brain activity, an elastic belt around the chest measures breathing effort, and a finger sensor measures blood oxygen level. The bottom panel (B) shows what the recorded data looks like over time. Each row represents a different measurement, including blood oxygen, breathing, and sleep stage. The bottom row tracks sleep stage, with the trace climbing upward during REM sleep and wakefulness and dropping downward during deep sleep. Adapted from PsychDB.
From Sleep Stages to Dream Conditions
Using polysomnography, researchers discovered that sleep is not a single uniform state [9]. Sleep cycles through distinct stages, each with a different brain signature. Deep sleep, sometimes called slow-wave sleep, produces large, slow brain waves and a calm, regular pattern. Another stage looks completely different. Brain activity becomes fast and almost wakelike, the eyes dart around behind closed lids in bursts, and most of the body’s voluntary muscles go limp, which is the brain’s way of keeping the dreamer from acting out whatever is happening internally. Researchers called this stage rapid eye movement sleep, or REM sleep. When sleepers were woken from REM and asked what they had been experiencing, they reported dreams far more often than people woken from deep sleep. That finding fixed REM in popular and scientific imagination as the stage of dreaming [9][11].
That early picture turned out to be too simple. People do report dreams from non-REM sleep, although those reports tend to be shorter, less vivid, and less emotional than REM reports. Neuroimaging complicated the picture further. Neuropsychologist Mark Solms reviewed cases in which damage to certain areas toward the front and middle of the brain reduced or abolished dreaming, even when REM sleep itself remained intact [12]. Furthermore, newer EEG studies that use many more electrodes than a standard sleep lab setup, giving a much sharper picture of which brain areas are active, identified a region toward the back of the brain whose activation predicted whether a sleeper would later report a dream, with the pattern holding in both REM and non-REM sleep [13]. These findings showed that REM and dreaming could come apart.
As a result, instead of asking in which sleep stage dreaming occurs, researchers shifted the question to ask under what conditions are dream experiences more likely or more intense. REM is one of the most reliable producers of those conditions. Sensory input from the world drops, voluntary movement is shut off, and subsequently, internally generated activity surges. For predictive processing, that matters because it suggests dreams are most vivid when the brain keeps generating a world-model while receiving less correction from the senses [1][12][13]. REM, therefore, is simply the most dramatic case in sleep of the brain’s model-building operating with the least sensory oversight.
Brainstem and Cortical Mechanisms of REM Dreaming
To understand more deeply how REM sleep creates the conditions for vivid dreaming, it is necessary to look at the mechanisms that produce the REM state. At a more detailed biological level, REM sleep depends on switching circuits in the brainstem, the stalk-like structure that connects the brain to the spinal cord. Researchers have identified groups of cells in the brainstem that turn REM on and other groups that turn it off. Nearby pathways also send signals that paralyze most of the body’s voluntary muscles during REM, which is why a person dreaming about running rarely actually runs. In this sense, the brainstem helps create the conditions for vivid dreaming, but it does not, by itself, explain why a dream contains vivid scenes, familiar people, or emotionally charged events. For that, we have to look at the cortex [14][15].
The cortex is the brain’s outer layer. It is involved in many of the processes that make conscious experience detailed and meaningful, including memory, language, planning, and interpretation. During REM sleep, several cortical systems shift in ways that help explain why dreams feel so real and so strange at the same time. First, visual construction remains active. Higher-order visual regions toward the back of the brain help build meaningful scenes rather than simply registering raw light. During REM, these regions remain active even though normal input from the eyes is reduced. The brain is still generating images, but the outside world is no longer tightly regulating what those images have to be. That is why dreams are not just replayed video clips from memory. They are active simulations assembled by a brain that is still building visual models from within.
Second, emotions are heightened. Activity rises in the amygdala, a small almond-shaped structure deep in the brain that helps tag experiences with emotional importance [16]. This helps explain why dream events can feel urgent, frightening, beautiful, or meaningful even when their plot makes little sense. The dream does not need to be logical to feel emotionally real. Third, reflective monitoring is weakened. Activity drops in frontal regions of the brain involved in planning and self-monitoring that would normally help us notice when something does not fit [17]. In a dream, a long-dead relative can appear in the kitchen, the kitchen can somehow also be a train station, and none of this strikes the dreamer as odd until morning. The dreamer often accepts what occurs in dreams without question because the systems that would usually flag the contradiction are less active. The result is a mental state that can be absurd and intense, while also oddly unquestioned [16][17][18].
This pattern fits the larger predictive processing story. In waking life, the brain’s model of the world is continually corrected by sensory input, bodily action, and reflective monitoring. In dreaming, those constraints loosen. Visual and emotional systems continue helping the brain build a world, while external correction and critical monitoring have less influence. The dream feels real not because it accurately represents the outside world, but because the brain is still using the same machinery that makes experience feel real in the first place.
What Dreams Reveal About Waking Perception
Ultimately, dreaming matters because it makes ordinary consciousness easier to see. In waking life, the brain’s world-model is held tightly in check by vision, touch, action, and the constant return of prediction errors from the environment and the body. The model never gets too far ahead of the world because the world keeps correcting it. In dreams, that same model-building capacity continues under radically altered conditions. External input is reduced, voluntary movement is suppressed, emotional and visual systems remain highly active, and the systems that catch contradictions go quiet. The result is not a completely different kind of mind, but the same mind with its balance changed. That is the deepest reason dreaming is worth studying. Every night the brain does what it always does, building a world from the inside out only now without the world watching over its shoulder. Dreams are thus amplified demonstrations of perception itself, nightly reminders of how the brain builds reality from within [1][2][7][13].
References
[1] Nir, Y., & Tononi, G. (2010). Dreaming and the brain: From phenomenology to neurophysiology. Trends in Cognitive Sciences, 14(2), 88–100. https://doi.org/10.1016/j.tics.2009.12.001
[2] Hobson, J. A., Hong, C. C.-H., & Friston, K. J. (2014). Virtual reality and consciousness inference in dreaming. Frontiers in Psychology, 5, Article 1133. https://doi.org/10.3389/fpsyg.2014.01133
[3] Revonsuo, A. (2000). The reinterpretation of dreams: An evolutionary hypothesis of the function of dreaming. Behavioral and Brain Sciences, 23(6), 877–901. https://doi.org/10.1017/S0140525X00004015
[4] Van der Helm, E., & Walker, M. P. (2009). Overnight therapy? The role of sleep in emotional brain processing. Psychological Bulletin, 135(5), 731–748. https://doi.org/10.1037/a0016570
[5] Wamsley, E. J., Tucker, M. A., Payne, J. D., Benavides, J. A., & Stickgold, R. (2010). Dreaming of a learning task is associated with enhanced sleep-dependent memory consolidation. Current Biology, 20(9), 850–855. https://doi.org/10.1016/j.cub.2010.03.027
[6] Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. Behavioral and Brain Sciences, 36(3), 181–204. https://doi.org/10.1017/S0140525X12000477
[7] Friston, K. (2010). The free-energy principle: A unified brain theory? Nature Reviews Neuroscience, 11(2), 127–138. https://doi.org/10.1038/nrn2787
[8] Velasco, P. F., & Loev, S. (2025). Metacognitive Feelings: A Predictive-Processing Perspective. Perspectives on Psychological Science. https://doi.org/10.1177/17456916231221976
[9] Aserinsky, E., & Kleitman, N. (1953). Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science, 118(3062), 273–274. https://doi.org/10.1126/science.118.3062.273
[10] Ruby, P. M. (2011). Experimental research on dreaming: State of the art and neuropsychoanalytic perspectives. Frontiers in Psychology, 2, Article 286. https://doi.org/10.3389/fpsyg.2011.00286
[11] Dement, W., & Kleitman, N. (1957). The relation of eye movements during sleep to dream activity: An objective method for the study of dreaming. Journal of Experimental Psychology, 53(5), 339–346. https://doi.org/10.1037/h0048189
[12] Solms, M. (2000). Dreaming and REM sleep are controlled by different brain mechanisms. Behavioral and Brain Sciences, 23(6), 843–850. https://doi.org/10.1017/S0140525X00003988
[13] Siclari, F., Baird, B., Perogamvros, L., Bernardi, G., LaRocque, J. J., Riedner, B., Boly, M., Postle, B. R., & Tononi, G. (2017). The neural correlates of dreaming. Nature Neuroscience, 20(6), 872–878. https://doi.org/10.1038/nn.4545
[14] Lu, J., Sherman, D., Devor, M., & Saper, C. B. (2006). A putative flip-flop switch for control of REM sleep. Nature, 441(7093), 589–594. https://doi.org/10.1038/nature04767
[15] Wehrle, R., Kaufmann, C., Wetter, T. C., Holsboer, F., Auer, D. P., Pollmächer, T., & Czisch, M. (2007). Functional microstates within human REM sleep: First evidence from fMRI of a thalamocortical network specific for phasic REM periods. European Journal of Neuroscience, 25(3), 863–871. https://doi.org/10.1111/j.1460-9568.2007.05314.x
[16] Maquet, P., Péters, J.-M., Aerts, J., Delfiore, G., Degueldre, C., Luxen, A., & Franck, G. (1996). Functional neuroanatomy of human rapid-eye-movement sleep and dreaming. Nature, 383(6596), 163–166. https://doi.org/10.1038/383163a0
[17] Braun, A. R., Balkin, T. J., Wesensten, N. J., Carson, R. E., Varga, M., Baldwin, P., Selbie, S., Belenky, G., & Herscovitch, P. (1997). Regional cerebral blood flow throughout the sleep-wake cycle. An H2(15)O PET study. Brain, 120(7), 1173–1197. https://doi.org/10.1093/brain/120.7.1173
[18] Braun, A. R., Balkin, T. J., Wesensten, N. J., Gwadry, F., Carson, R. E., Varga, M., Baldwin, P., Belenky, G., & Herscovitch, P. (1998). Dissociated pattern of activity in visual cortices and their projections during human rapid eye movement sleep. Science, 279(5347), 91–95. https://doi.org/10.1126/science.279.5347.91